GPCRを介する治療薬に対するバイアスの詳細な機序は未解明だった
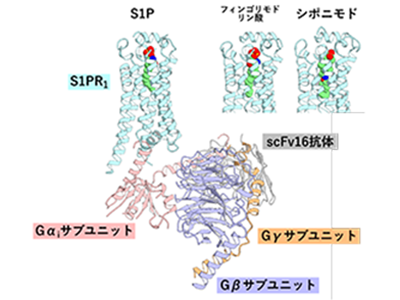
東北大学は12月24日、3種類の化合物(S1P、フィンゴリモドの代謝活性体、シポニモド)が、それぞれ結合したS1PR1の立体構造を解明し、それに基づいた分子動力学シミュレーションと機能解析を行うことで、治療薬結合型受容体の作動基盤を解明したと発表した。この研究は、同大大学院薬学研究科の井上飛鳥准教授らの研究グループと、中国ハルビン工業大学のYuanzheng He教授らのグループとの共同研究によるもの。研究成果は、「Nature Chemical Biology」の電子版に掲載されている。
画像はリリースより
スフィンゴ脂質の一種であるS1Pは、前駆体であるスフィンゴシンが細胞内でスフィンゴシンキナーゼによってリン酸化されることで生合成される。S1Pは輸送体によって細胞外に放出されると、近傍の細胞の表面に存在するGタンパク質共役受容体(GPCR)の一種であるS1P受容体に結合・活性化することで、下流シグナル因子である三量体Gタンパク質のシグナル伝達を誘導する。S1P受容体は5種類(S1PR1-S1PR5)存在するが、なかでもS1PR1は免疫に関わるTリンパ球に高発現していることが知られている。Tリンパ球は、血管とリンパ管を循環して体内の異物を監視しており、リンパ節から血中への移行の際には、S1Pの濃度勾配によるTリンパ球の遊走が必須であることが知られている。血中の高濃度のS1Pに晒されるとTリンパ球のS1PR1は細胞内へと内在化し、S1P濃度の低いリンパ管に移行して、S1PR1が再び細胞表面に出てくるまでS1Pに不応答となる。免疫抑制剤として開発された薬剤であるフィンゴリモドは、体内に取り込まれるとS1Pと同様に、スフィンゴシンキナーゼによって代謝活性体であるリン酸化体に変換される。このフィンゴリモドリン酸はS1Pと同様にS1PR1に結合し(結合分子を一般にリガンドと呼ぶ)、三量体Gタンパク質のシグナル伝達を誘導する。
一方で、フィンゴリモドリン酸はS1PR1をS1Pよりも強く内在化させ、最終的に細胞内で分解させることによって、Tリンパ球の体内循環を阻害して免疫を抑制すると考えられている。S1PR1を含むGPCRはアレスチンと呼ばれるシグナル調節タンパク質と結合することで内在化されることが知られている。フィンゴリモドリン酸が結合したS1PR1は、三量体Gタンパク質よりもアレスチンを好むことから、アレスチンバイアスリガンド(アレスチンに偏る結合因子の意味)と呼ばれている。また、2020年に日本で多発性硬化症の治療薬として認可されたシポニモドもアレスチンバイアスリガンドであることが知られている。しかし、これらの治療薬がなぜバイアスを引き起こすのかは不明なままであり、GPCR一般に対するバイアスの詳細な機序も解明されていなかった。
多発性硬化症治療薬結合時に特徴的なS1PR1内部の構造変化を発見
今回、研究グループは、S1Pやフィンゴリモドリン酸、シポニモドが結合したS1PR1および三量体Gタンパク質で構成されるシグナル伝達複合体の立体構造を、クライオ電子顕微鏡を用いて決定した。そして、これらの構造に基づいたS1PR1の1アミノ酸変異体の機能解析により、リガンドの認識機構を明らかにした。また、これらの構造に基づく分子動力学シミュレーションにより、アレスチンバイアスリガンドに特有のS1PR1の内部の構造変化を見出した。さらに、特定された構造変化に関わるアミノ酸残基を置換したS1PR1変異体を作製し、三量体Gタンパク質の活性化とS1PR1とアレスチン複合体の内在化を測定することで、見出された構造変化がバイアスに関わることを実証した。
F265とN307の相互作用がS1PR1機能のバイアス性の起因だった
GPCRはリガンドが細胞外から結合すると、その立体構造が変化し、細胞内にGα/Gβ/Gγの3つのサブユニットからなる三量体Gタンパク質を結合する。その後、三量体Gタンパク質は核酸交換反応を経て、GαサブユニットとGβγサブユニットに解離する。研究グループは、三量体Gタンパク質の改変体を用いてこの解離を測定し、S1PR1のGタンパク質経路を測定した。S1PR1とともに分割ルシフェラーゼを融合したGαサブユニットとGγサブユニットを培養細胞に発現させると、ルシフェラーゼ由来の発光が検出できる。この状態で、S1PR1リガンドを添加してGタンパク質が活性化されると、発光の減弱を継時的に測定できる。この評価系を用いて、種々の変異を導入したS1PR1の三量体Gタンパク質シグナルを評価した。リガンド結合ポケットのアミノ酸残基を変異させて(120番目のアルギニン(R120)と121番目のグルタミン酸(E121)をそれぞれアラニンに置換)、三量体Gタンパク質の活性化を測定したところ、S1Pとフィンゴリモドリン酸は類似の認識機構によってS1PR1に結合する一方、シポニモドは別のアミノ酸残基を介した認識機構によってS1PR1に結合することがわかった。
これら3種のリガンドが結合したS1PR1のクライオ電顕構造は、全体として極めて似通った構造をとっており、バイアス性がどのように生じるか不明だった。そこで、分子動力学シミュレーションと呼ばれる計算機シミュレーションを行い、運動方程式に従った原子の経時変化を追跡。このシミュレーションを異なるリガンドが結合した状態のS1PR1に対して行い、それぞれの結果を解析・比較した。その結果、アレスチンバイアスリガンドであるフィンゴリモドリン酸とシポニモドに共通し、S1Pとは異なるS1PR1の内部のアミノ酸残基同士の相互作用を見出した。そこで、この残基を変異させたS1PR1がどのようなバイアスを示すのかについて実験で確かめた。上述の三量体Gタンパク質経路に関する測定に加え、細胞膜やエンドソームにおけるアレスチンとS1PR1の複合体の局在をルシフェラーゼ断片の相互作用によって評価。その結果、着目した相互作用(265番目のフェニルアラニン(F265)と307番目のアスパラギン(N307))を低下させたアラニン変異体では、三量体Gタンパク質とアレスチンのバランスが変化することを確認でき、これらの相互作用がS1PR1機能のバイアス性の起因であることを明らかにした。
S1P受容体に限らず、他のGPCRに対しても適用できる可能性
今回の研究は、S1PR1に対する薬剤の結合様式を示し、今後のS1PR1に対する薬剤開発に役立つと期待される。また、アレスチンバイアスリガンド特有の受容体内部の相互作用はGPCRによく保存された残基が関わるため、S1P受容体に限らず、他の多くのGPCRに対しても適用可能であることが期待される、と研究グループは述べている。
▼関連リンク
・東北大学 プレスリリース

